
Veškerý obsah iLive je lékařsky zkontrolován nebo zkontrolován, aby byla zajištěna co největší věcná přesnost.
Máme přísné pokyny pro získávání zdrojů a pouze odkaz na seriózní mediální stránky, akademické výzkumné instituce a, kdykoli je to možné, i klinicky ověřené studie. Všimněte si, že čísla v závorkách ([1], [2] atd.) Jsou odkazy na tyto studie, na které lze kliknout.
Pokud máte pocit, že některý z našich obsahů je nepřesný, neaktuální nebo jinak sporný, vyberte jej a stiskněte klávesu Ctrl + Enter.
hypofýza
Lékařský expert článku

Naposledy posuzováno: 04.07.2025

Hypofýza (hypofýza, s.glandula pituitaria) se nachází v hypofyzární jámě turecké sella sfenoidní kosti a od lebeční dutiny je oddělena výběžkem tvrdé pleny mozkové, tvořícím bránici sella. Otvorem v této bránici je hypofýza spojena s infundibulem hypotalamu diencephalonu. Příčný rozměr hypofýzy je 10-17 mm, předozadní 5-15 mm, vertikální 5-10 mm. Hmotnost hypofýzy u mužů je přibližně 0,5 g, u žen 0,6 g. Hypofýza je zvenčí pokryta pouzdrem.
V souladu s vývojem hypofýzy ze dvou různých zárodků se v orgánu rozlišují dva laloky - přední a zadní. Adenohypofýza neboli přední lalok (adenohypofýza, s.lobus anterior) je větší a tvoří 70-80 % celkové hmotnosti hypofýzy. Je hustší než zadní lalok. V předním laloku se rozlišuje distální část (pars distalis), která zabírá přední část hypofyzární jamky, střední část (pars intermedia), která se nachází na hranici se zadním lalokem, a hlíznatá část (pars tuberalis), která směřuje vzhůru a spojuje se s infundibulem hypotalamu. Vzhledem k hojnosti krevních cév má přední lalok světle žlutou barvu s načervenalým odstínem. Parenchym přední hypofýzy je reprezentován několika typy žlázových buněk, mezi jejichž prameny se nacházejí sinusoidní krevní kapiláry. Polovina (50 %) buněk adenohypofýzy jsou chromofilní adenocyty, které mají ve své cytoplazmě jemnozrnné granule, jež se dobře barví solemi chromu. Jedná se o acidofilní adenocyty (40 % všech buněk adenohypofýzy) a bazofilní adenocyty (10 %). Mezi bazofilní adenocyty patří gonadotropní, kortikotropní a tyreotropní endokrinocyty. Chromofobní adenocyty jsou malé, mají velké jádro a malé množství cytoplazmy. Tyto buňky jsou považovány za prekurzory chromofilních adenocytů. Zbývajících 50 % buněk adenohypofýzy jsou chromofobní adenocyty.
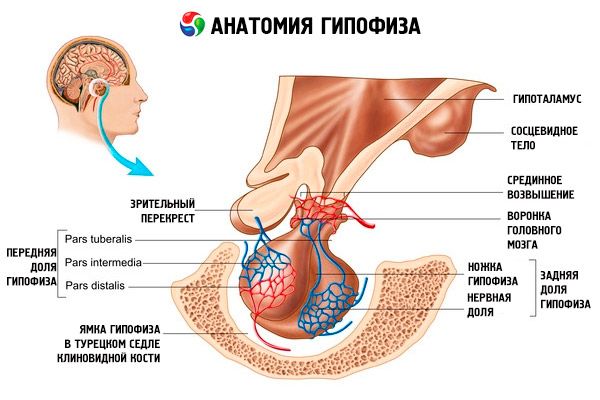
Neurohypofýza neboli zadní lalok (neurohypophysis, s.lobus posterior) se skládá z nervového laloku (lobus nervosus), který se nachází v zadní části hypofyzární jamky, a trychtýře (infundibulum), který se nachází za hlíznou částí adenohypofýzy. Zadní lalok hypofýzy je tvořen neurogliálními buňkami (buňkami hypofýzy), nervovými vlákny, která jdou z neurosekrečních jader hypotalamu do neurohypofýzy, a neurosekrečními tělísky.
Hypofýza je prostřednictvím nervových vláken (drážek) a cév funkčně propojena s hypotalamem (mezimozkem), který reguluje činnost hypofýzy. Hypofýza a hypotalamus se spolu s jejich neuroendokrinními, cévními a nervovými spojeními obvykle považují za hypotalamo-hypofyzární systém.
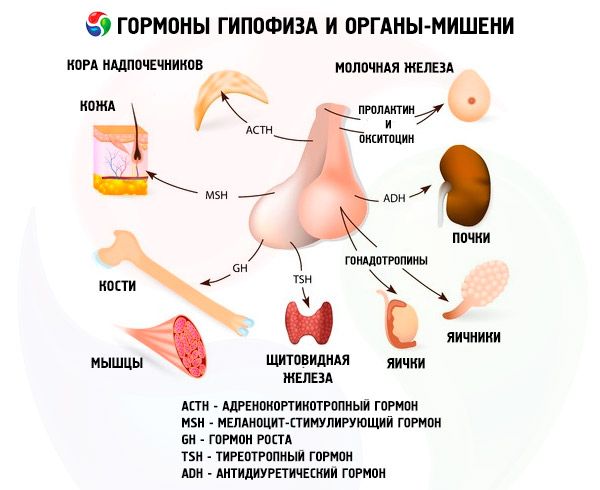
Hormony přední a zadní hypofýzy ovlivňují mnoho funkcí těla, především prostřednictvím jiných endokrinních žláz. V přední hypofýze produkují acidofilní adenocyty (alfa buňky) somatotropní hormon (růstový hormon), který se podílí na regulaci růstových a vývojových procesů u mladých organismů. Kortikotropní endokrinocyty vylučují adrenokortikotropní hormon (ACTH), který stimuluje sekreci steroidních hormonů nadledvinami. Tyreotropní endokrinocyty vylučují tyreotropní hormon (TSH), který ovlivňuje vývoj štítné žlázy a aktivuje produkci jejích hormonů. Gonadotropní hormony: folikulostimulační (FSH), luteinizační (LH) a prolaktin - ovlivňují pohlavní zrání těla, regulují a stimulují vývoj folikulů ve vaječníku, ovulaci, růst mléčných žláz a produkci mléka u žen, proces spermatogeneze u mužů. Tyto hormony produkují bazofilní adenocyty (beta buňky). Zde se vylučují i lipotropní faktory hypofýzy, které ovlivňují mobilizaci a využití tuků v těle. V mezilehlé části předního laloku se tvoří melanocyty stimulující hormon, který v těle řídí tvorbu pigmentů - melaninů.
Neurosekreční buňky supraoptických a paraventrikulárních jader v hypotalamu produkují vasopresin a oxytocin. Tyto hormony jsou transportovány do buněk zadní hypofýzy podél axonů, které tvoří hypotalamo-hypofyzární trakt. Ze zadní hypofýzy se tyto látky dostávají do krve. Hormon vasopresin má vazokonstrikční a antidiuretický účinek, pro který se také nazývá antidiuretickým hormonem (ADH). Oxytocin má stimulační účinek na kontraktilitu děložních svalů, zvyšuje sekreci mléka kojící mléčnou žlázou, inhibuje vývoj a funkci žlutého tělíska a ovlivňuje změny tónu hladkých (nepruhovaných) svalů gastrointestinálního traktu.
Vývoj hypofýzy
Přední lalok hypofýzy se vyvíjí z epitelu dorzální stěny ústní díry ve formě prstencovitého výrůstku (Rathkeho váček). Tento ektodermální výběžek roste směrem ke dnu budoucí třetí mozkové komory. Směrem k němu, ze spodního povrchu druhého mozkového váčku (budoucí dno třetí komory), vyrůstá výběžek, ze kterého se vyvíjí šedý tuberkul infundibula a zadní lalok hypofýzy.
Cévy a nervy hypofýzy
Z vnitřních krčních tepen a cév arteriálního kruhu mozku směřují horní a dolní hypofyzární tepny do hypofýzy. Horní hypofyzární tepny jdou do šedého jádra a infundibula hypotalamu, zde se vzájemně anastomují a tvoří kapiláry pronikající mozkovou tkání - primární hemokapilární síť. Z dlouhých a krátkých smyček této sítě se tvoří portální žíly, které směřují do předního laloku hypofýzy. V parenchymu předního laloku hypofýzy se tyto žíly rozpadají na široké sinusoidní kapiláry a tvoří sekundární hemokapilární síť. Zadní lalok hypofýzy je zásoben krví převážně dolní hypofyzární tepnou. Mezi horní a dolní hypofyzární tepnou jsou dlouhé arteriální anastomózy. Odtok žilní krve ze sekundární hemokapilární sítě se uskutečňuje systémem žil, které ústí do kavernózních a interkavernózních sinusů dura mater mozku.
Sympatická vlákna, která vstupují do orgánu spolu s tepnami, se podílejí na inervaci hypofýzy. Postgangliová sympatická nervová vlákna odcházejí z plexu vnitřní karotické tepny. Kromě toho se v zadním laloku hypofýzy nacházejí četné zakončení výběžků neurosekrečních buněk umístěných v jádrech hypotalamu.
Věkové rysy hypofýzy
Průměrná hmotnost hypofýzy u novorozenců dosahuje 0,12 g. Hmotnost orgánu se do 10 let věku zdvojnásobí a do 15 let ztrojnásobí. Do 20 let věku dosahuje hmotnost hypofýzy svého maxima (530-560 mg) a v následujících věkových obdobích zůstává téměř nezměněna. Po 60 letech je pozorován mírný pokles hmotnosti této endokrinní žlázy.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hormony hypofýzy
Jednota nervové a hormonální regulace v těle je zajištěna úzkým anatomickým a funkčním propojením hypofýzy a hypotalamu. Tento komplex určuje stav a fungování celého endokrinního systému.
Hlavní endokrinní žlázou, která produkuje řadu peptidových hormonů přímo regulujících funkci periferních žláz, je hypofýza. Jedná se o červenošedý útvar ve tvaru fazole pokrytý vláknitou kapslí o hmotnosti 0,5-0,6 g. Jeho velikost se mírně liší v závislosti na pohlaví a věku osoby. Obecně přijímané rozdělení hypofýzy na dva laloky, které se liší vývojem, strukturou a funkcí: přední distální - adenohypofýza a zadní - neurohypofýza. První tvoří asi 70 % celkové hmotnosti žlázy a je konvenčně rozdělen na distální, infundibulární a střední část, druhý - na zadní část neboli lalok a stopku hypofýzy. Žláza se nachází v hypofyzární jamce turckého sedla klínové kosti a je stopkou spojena s mozkem. Horní část předního laloku je pokryta optickým chiasmatem a optickými dráhami. Krevní zásobení hypofýzy je velmi hojné a zajišťuje se větvemi vnitřní karotidy (horní a dolní hypofyzární tepny) a také větvemi arteriálního kruhu mozku. Horní hypofyzární tepny se podílejí na krevním zásobení adenohypofýzy a dolní neurohypofýzy, přičemž se dotýkají neurosekretorických zakončení axonů velkých buněčných jader hypotalamu. První z nich vstupují do střední eminence hypotalamu, kde se rozptylují do kapilární sítě (primární kapilární plexus). Tyto kapiláry (s nimiž se dotýkají zakončení axonů malých neurosekretorických buněk mediobazálního hypotalamu) se shromažďují do portálních žil sestupujících podél stopky hypofýzy do parenchymu adenohypofýzy, kde se opět dělí do sítě sinusoidních kapilár (sekundární kapilární plexus). Krev tedy po předchozím průchodu střední eminencí hypotalamu, kde je obohacena hypotalamickými adenohypofyzotropními hormony (uvolňujícími hormony), vstupuje do adenohypofýzy.
Odtok krve nasycené hormony adenohypofýzy z četných kapilár sekundárního plexu se provádí systémem žil, které zase ústí do žilních sinusů dura mater a poté do celkového krevního oběhu. Portální systém hypofýzy se sestupným směrem toku krve z hypotalamu je tedy morfofunkční složkou komplexního mechanismu neurohumorální regulace tropických funkcí adenohypofýzy.
Hypofýza je inervována sympatickými vlákny, která navazují na hypofyzární tepny. Vycházejí z postgangliových vláken, která procházejí vnitřním karotickým plexem a jsou spojena s horními cervikálními ganglii. Adenohypofýza není přímo inervována hypothalamem. Zadní lalok přijímá nervová vlákna z neurosekrečních jader hypotalamu.
Adenohypofýza je co do histologické architektury velmi složitý útvar. Má dva typy žlázových buněk - chromofobní a chromofilní. Ty se zase dělí na acidofilní a bazofilní (podrobný histologický popis hypofýzy je uveden v odpovídající části manuálu). Je však třeba poznamenat, že hormony produkované žlázovými buňkami, které tvoří parenchym adenohypofýzy, se vzhledem k jejich rozmanitosti do jisté míry liší svou chemickou povahou a jemná struktura sekretujících buněk musí odpovídat biosyntetickým vlastnostem každé z nich. Někdy však lze v adenohypofýze pozorovat přechodné formy žlázových buněk, které jsou schopny produkovat několik hormonů. Existují důkazy o tom, že typ žlázových buněk adenohypofýzy není vždy určen geneticky.
Pod bránicí turecké sedla se nachází trychtýřovitá část předního laloku. Objímá stopku hypofýzy a dotýká se šedého tuberkulu. Tato část adenohypofýzy se vyznačuje přítomností epiteliálních buněk a bohatým prokrvením. Je také hormonálně aktivní.
Střední (mezilehlá) část hypofýzy se skládá z několika vrstev velkých sekrečně aktivních bazofilních buněk.
Hypofýza plní prostřednictvím svých hormonů různé funkce. Její přední lalok produkuje adrenokortikotropní (ACTH), tyreostimulační (TSH), folikulostimulační (FSH), luteinizační (LH) a lipotropní hormony, a také růstový hormon - somatotropní (STO) a prolaktin. V mezilehlém laloku se syntetizuje melanocyty stimulující hormon (MSH) a v zadním laloku se hromadí vasopresin a oxytocin.
ACTH
Hypofyzární hormony jsou skupinou proteinových a peptidových hormonů a glykoproteinů. Z hormonů přední hypofýzy je ACTH nejlépe prozkoumán. Je produkován bazofilními buňkami. Jeho hlavní fyziologickou funkcí je stimulace biosyntézy a sekrece steroidních hormonů kůrou nadledvin. ACTH také vykazuje melanocyty stimulující a lipotropní aktivitu. V roce 1953 byl izolován v čisté formě. Později byla stanovena jeho chemická struktura, která se u lidí a řady savců skládá z 39 aminokyselinových zbytků. ACTH nemá druhovou specificitu. V současné době byla provedena chemická syntéza jak samotného hormonu, tak různých fragmentů jeho molekuly, aktivnějších než přirozené hormony. Struktura hormonu má dvě části peptidového řetězce, z nichž jedna zajišťuje detekci a vazbu ACTH na receptor a druhá poskytuje biologický účinek. Zřejmě se váže na receptor ACTH interakcí elektrických nábojů hormonu a receptoru. Úlohu biologického efektoru ACTH plní fragment molekuly 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
Melanocyty stimulující aktivita ACTH je způsobena přítomností N-terminální oblasti v molekule, která se skládá ze 13 aminokyselinových zbytků a opakuje strukturu alfa-melanocyty stimulujícího hormonu. Tato oblast také obsahuje heptapeptid přítomný v jiných hypofyzárních hormonech a vykazuje určité adrenokortikotropní, melanocyty stimulující a lipotropní účinky.
Za klíčový moment v působení ACTH se považuje aktivace enzymu proteinkinázy v cytoplazmě za účasti cAMP. Fosforylovaná proteinkináza aktivuje enzym esterázu, která přeměňuje estery cholesterolu na volnou látku v tukových kapénkách. Protein syntetizovaný v cytoplazmě v důsledku fosforylace ribozomů stimuluje vazbu volného cholesterolu na cytochrom P-450 a jeho přenos z lipidových kapének do mitochondrií, kde jsou přítomny všechny enzymy zajišťující přeměnu cholesterolu na kortikosteroidy.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Hormon stimulující štítnou žlázu
TSH - tyreotropin - hlavní regulátor vývoje a funkce štítné žlázy, procesů syntézy a sekrece hormonů štítné žlázy. Tento komplexní protein - glykoprotein - se skládá z alfa a beta podjednotek. Struktura první podjednotky se shoduje s alfa podjednotkou luteinizačního hormonu. Navíc se do značné míry shoduje u různých druhů zvířat. Sekvence aminokyselinových zbytků v beta podjednotce lidského TSH byla dešifrována a skládá se ze 119 aminokyselinových zbytků. Lze poznamenat, že beta podjednotky lidského a hovězího TSH jsou si v mnoha ohledech podobné. Biologické vlastnosti a povaha biologické aktivity glykoproteinových hormonů jsou určeny beta podjednotkou. Ta také zajišťuje interakci hormonu s receptory v různých cílových orgánech. Beta podjednotka však u většiny zvířat vykazuje specifickou aktivitu až po spojení s alfa podjednotkou, která působí jako druh aktivátoru hormonu. Ta se stejnou pravděpodobností indukuje luteinizační, folikulostimulační a tyreotropní aktivity určené vlastnostmi beta podjednotky. Zjištěná podobnost nám umožňuje dojít k závěru, že tyto hormony vznikly v procesu evoluce z jednoho společného předchůdce, přičemž beta podjednotka také určuje imunologické vlastnosti hormonů. Existuje předpoklad, že alfa podjednotka chrání beta podjednotku před působením proteolytických enzymů a také usnadňuje její transport z hypofýzy do periferních cílových orgánů.
Gonadotropní hormony
Gonadotropiny jsou v těle přítomny ve formě LH a FSH. Funkční účel těchto hormonů se obecně omezuje na zajištění reprodukčních procesů u jedinců obou pohlaví. Stejně jako TSH jsou to komplexní proteiny - glykoproteiny. FSH indukuje zrání folikulů ve vaječnících žen a stimuluje spermatogenezi u mužů. LH způsobuje u žen prasknutí folikulů s tvorbou žlutého tělíska a stimuluje sekreci estrogenů a progesteronu. U mužů stejný hormon urychluje vývoj intersticiální tkáně a sekreci androgenů. Účinky gonadotropinů jsou na sobě závislé a probíhají synchronně.
Dynamika sekrece gonadotropinů u žen se během menstruačního cyklu mění a byla dostatečně podrobně studována. V preovulační (folikulární) fázi cyklu je obsah LH na poměrně nízké úrovni, zatímco FSH je zvýšený. S dozráváním folikulu se zvyšuje sekrece estradiolu, což podporuje zvýšenou produkci gonadotropinů hypofýzou a výskyt cyklů LH i FSH, tj. pohlavní steroidy stimulují sekreci gonadotropinů.
V současné době byla struktura LH stanovena. Stejně jako TSH se skládá ze 2 podjednotek: a a b. Struktura alfa podjednotky LH se u různých druhů zvířat do značné míry shoduje, odpovídá struktuře alfa podjednotky TSH.
Struktura beta podjednotky LH se významně liší od struktury beta podjednotky TSH, ačkoli má čtyři identické úseky peptidového řetězce sestávající ze 4-5 aminokyselinových zbytků. V TSH jsou lokalizovány v pozicích 27-31, 51-54, 65-68 a 78-83. Vzhledem k tomu, že beta podjednotka LH a TSH určuje specifickou biologickou aktivitu hormonů, lze předpokládat, že homologní úseky ve struktuře LH a TSH by měly zajišťovat spojení beta podjednotek s alfa podjednotkou a úseky, které se liší strukturou, by měly být zodpovědné za specifičnost biologické aktivity hormonů.
Nativní LH je velmi stabilní vůči působení proteolytických enzymů, ale beta podjednotka je chymotrypsinem rychle štěpena a podjednotka a je enzymem obtížně hydrolyzovatelná, tj. hraje ochrannou roli a brání chymotrypsinu v přístupu k peptidovým vazbám.
Pokud jde o chemickou strukturu FSH, vědci dosud nezískali definitivní výsledky. Stejně jako LH se FSH skládá ze dvou podjednotek, ale beta podjednotka FSH se liší od beta podjednotky LH.
Prolaktin
Další hormon, prolaktin (laktogenní hormon), hraje aktivní roli v reprodukčních procesech. Hlavní fyziologické vlastnosti prolaktinu u savců se projevují ve formě stimulace vývoje mléčných žláz a laktace, růstu mazových žláz a vnitřních orgánů. Podporuje projevy účinku steroidů na sekundární pohlavní znaky u samců, stimuluje sekreční aktivitu žlutého tělíska u myší a potkanů a podílí se na regulaci metabolismu tuků. V posledních letech je prolaktinu věnována velká pozornost jako regulátoru mateřského chování; tato polyfunkčnost je vysvětlena jeho evolučním vývojem. Je to jeden z dávných hormonů hypofýzy a nachází se i u obojživelníků. V současné době je struktura prolaktinu u některých druhů savců zcela rozluštěna. Až donedávna však vědci vyjadřovali pochybnosti o existenci takového hormonu u lidí. Mnozí se domnívali, že jeho funkci plní růstový hormon. Nyní byly získány přesvědčivé důkazy o přítomnosti prolaktinu u lidí a jeho struktura byla částečně rozluštěna. Prolaktinové receptory aktivně vážou růstový hormon a placentární laktogen, což naznačuje jediný mechanismus účinku těchto tří hormonů.
Somatotropin
Růstový hormon, somatotropin, má ještě širší spektrum účinku než prolaktin. Stejně jako prolaktin je produkován acidofilními buňkami adenohypofýzy. STH stimuluje růst kostry, aktivuje biosyntézu bílkovin, má účinek mobilizace tuků a podporuje zvětšení tělesné velikosti. Kromě toho koordinuje metabolické procesy.
Účast hormonu v tomto druhém případě je potvrzena skutečností prudkého zvýšení jeho sekrece hypofýzou, například při poklesu hladiny cukru v krvi.
Chemická struktura tohoto lidského hormonu je nyní plně objasněna – 191 aminokyselinových zbytků. Jeho primární struktura je podobná struktuře choriového somatomammotropinu nebo placentárního laktogenu. Tato data naznačují významnou evoluční blízkost těchto dvou hormonů, ačkoli vykazují rozdíly v biologické aktivitě.
Je nutné zdůraznit vysokou druhovou specificitu daného hormonu - například STH živočišného původu je u lidí neaktivní. To se vysvětluje jak reakcí mezi receptory STH lidí a zvířat, tak i strukturou samotného hormonu. V současné době probíhají studie zaměřené na identifikaci aktivních center ve složité struktuře STH, která vykazují biologickou aktivitu. Studují se jednotlivé fragmenty molekuly, které vykazují další vlastnosti. Například po hydrolýze lidského STH pepsinem byl izolován peptid sestávající ze 14 aminokyselinových zbytků a odpovídající úseku molekuly 31-44. Neměl růstový účinek, ale v lipotropní aktivitě významně převyšoval nativní hormon. Lidský růstový hormon má na rozdíl od podobného hormonu zvířat významnou laktogenní aktivitu.
Adenohypofýza syntetizuje mnoho peptidových a proteinových látek, které mají tuk-mobilizující účinek, a lipotropní účinek mají hypofyzární tropní hormony - ACTH, STH, TSH a další. V posledních letech se zvláště zdůrazňují beta- a γ-lipotropní hormony (LPG). Biologické vlastnosti beta-LPG byly studovány nejpodrobněji; kromě lipotropní aktivity má také melanocyty stimulující, kortikotropin stimulující a hypokalcemický účinek a také vyvolává inzulínu podobný účinek.
V současné době byla dešifrována primární struktura ovčího LPG (90 aminokyselinových zbytků), lipotropních hormonů prasat a skotu. Tento hormon má druhovou specificitu, ačkoli struktura centrální oblasti beta-LPG je u různých druhů stejná. To určuje biologické vlastnosti hormonu. Jeden z fragmentů této oblasti se nachází ve struktuře alfa-MSH, beta-MSH, ACTH a beta-LPG. Předpokládá se, že tyto hormony vznikly v procesu evoluce ze stejného prekurzoru. γ-LPG má slabší lipotropní aktivitu než beta-LPG.
Melanocyty stimulující hormon
Tento hormon, syntetizovaný v mezilehlém laloku hypofýzy, stimuluje biosyntézu kožního pigmentu melaninu v jeho biologické funkci, podporuje zvětšení velikosti a počtu pigmentových buněk melanocytů v kůži obojživelníků. Tyto vlastnosti MSH se používají při biologickém testování hormonu. Existují dva typy hormonu: alfa- a beta-MSH. Bylo prokázáno, že alfa-MSH nemá druhovou specificitu a má stejnou chemickou strukturu u všech savců. Jeho molekula je peptidový řetězec sestávající ze 13 aminokyselinových zbytků. Beta-MSH má naopak druhovou specificitu a jeho struktura se u různých zvířat liší. U většiny savců se molekula beta-MSH skládá z 18 aminokyselinových zbytků a pouze u lidí je od aminokyselinového konce prodloužena o čtyři aminokyselinové zbytky. Je třeba poznamenat, že alfa-MSH má určitou adrenokortikotropní aktivitu a jeho vliv na chování zvířat i lidí byl nyní prokázán.
Oxytocin a vasopresin
Vasopresin a oxytocin, které se syntetizují v hypotalamu, se hromadí v zadním laloku hypofýzy: vasopresin v neuronech supraoptického jádra a oxytocin v paraventrikulačním jádru. Poté se přenášejí do hypofýzy. Je třeba zdůraznit, že prekurzor hormonu vasopresinu se syntetizuje nejprve v hypotalamu. Současně se zde produkují proteiny neurofyzinů typu 1 a 2. První se váže na oxytocin a druhý na vasopresin. Tyto komplexy migrují ve formě neurosekrečních granulí v cytoplazmě podél axonu a dosahují zadního laloku hypofýzy, kde nervová vlákna končí v cévní stěně a obsah granulí vstupuje do krve. Vasopresin a oxytocin jsou prvními hormony hypofýzy s plně stanovenou aminokyselinovou sekvencí. Ve své chemické struktuře se jedná o nonapeptidy s jedním disulfidovým můstkem.
Uvažované hormony vyvolávají různé biologické účinky: stimulují transport vody a solí membránami, mají vazopresorický účinek, zvyšují kontrakce hladkého svalstva dělohy během porodu a zvyšují sekreci mléčných žláz. Je třeba poznamenat, že vazopresin má vyšší antidiuretickou aktivitu než oxytocin, zatímco oxytocin má silnější účinek na dělohu a mléčnou žlázu. Hlavním regulátorem sekrece vazopresinu je spotřeba vody; v renálních tubulech se váže na receptory v cytoplazmatických membránách s následnou aktivací enzymu adenylátcyklázy. Za vazbu hormonu na receptor a za biologický účinek jsou zodpovědné různé části molekuly.
Hypofýza, propojená přes hypotalamus s celým nervovým systémem, spojuje endokrinní systém do funkčního celku a podílí se na zajištění stálosti vnitřního prostředí těla (homeostáza). V rámci endokrinního systému se homeostatická regulace provádí na principu zpětné vazby mezi předním lalokem hypofýzy a „cílovými“ žlázami (štítná žláza, kůra nadledvin, pohlavní žlázy). Nadbytek hormonu produkovaného „cílovou“ žlázou inhibuje a jeho nedostatek stimuluje sekreci a uvolňování odpovídajícího tropního hormonu. Hypotalamus je součástí systému zpětné vazby. Právě v něm se nacházejí receptorové zóny citlivé na hormony „cílových“ žláz. Specifickou vazbou na hormony cirkulující v krvi a změnou reakce v závislosti na koncentraci hormonů přenášejí hypotalamické receptory svůj účinek do odpovídajících hypotalamických center, která koordinují práci adenohypofýzy a uvolňují hypotalamické adenohypofyziotropní hormony. Hypotalamus by tedy měl být považován za neuroendokrinní mozek.
Использованная литература